Qu’est-ce que l’ouïe ?

L’ouïe est le terme utilisé pour désigner le sens auquel correspond l’action d’entendre : c’est l’audition, le rôle de l’oreille.
Les 3 grandes parties de l’oreilles : anatomie
Image 1
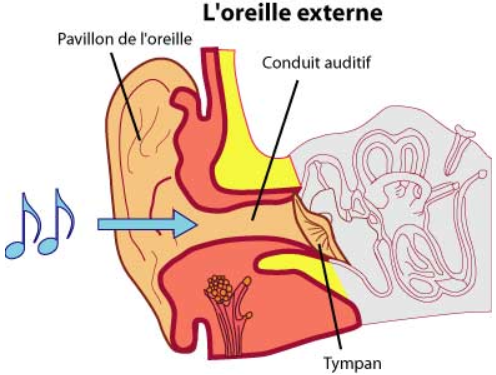
- Oreille externe : Elle est constituée du conduit auditif externe (CAE), et du tympan et permet l’entrée des ondes sonores. C’est le pavillon qui capte les ondes sonores (vibrations de l’air) et les transmet au tympan via le conduit auditif, ensuite on atteint l’oreille moyenne.
Image 2

- Oreille moyenne : cavité remplie d’air qui abrite la caisse du tympan, la trompe d’Eustache et la chaîne des osselets. On y trouve les trois os les plus petits du corps humain : le marteau qui est au contact du tympan, l’enclume, et l’étrier. Ils sont reliés d’un côté au tympan et de l’autre à l’oreille interne par une fine membrane. Leur rôle est d’amplifier la vibration captée par le tympan. Cette chaîne osseuse dont les éléments ont une taille de l’ordre du millimètre réalise la jonction entre l’oreille externe et l’oreille interne. La fonction principale de la trompe d’Eustache est d’égaliser la pression de l’air dans l’oreille moyenne avec celle de l’air ambiant, elle met l’oreille moyenne en communication avec la gorge. Lorsque l’on est pris de la gorge, ce canal peut parfois rester bouché, ce qui s’accompagne d’une atténuation passagère de l’audition.
Image 3

- Oreille interne : composée de la cochlée, du vestibule et des canaux semi-circulaires. Cette partie de l’oreille est reliée à l’oreille moyenne par deux orifices : la fenêtre ovale et la fenêtre ronde qui sont obturées par des membranes. L’étrier est un os qui donne sur la cochlée et qui s’insère sur la fenêtre ovale. La cochlée est responsable du traitement du signal et les canaux semi-circulaires, de l’équilibre. Le canal cochléaire, rempli de liquide, est muni de fines cellules ciliées sur toute sa longueur.
Dans l’oreille interne se trouve un organe sensoriel barosensible qui ne joue pas de rôle particulier dans l’audition : c’est le système vestibulaire. Il contribue à la sensation de mouvement et à l’équilibre chez la plupart des mammifères. C’est pourquoi nous ne le traiterons pas sur notre site.
Tous ces éléments ne sont pas indispensable pour l’audition, nous traiterons ceux qui permettre d’entendre dans les parties suivantes.
LE SAVIEZ-VOUS ?
Notre acuité visuelle diminue au fur et à mesure du temps : c’est la presbytie, il en va de même pour l’audition : c’est ce que l’on appelle la presbyacousie. Cela correspond au vieillissement normal de l’oreille interne qui débute dès 25 ans et qui se traduit par une perte progressive de l’audition avec l’âge. C’est la cause la plus fréquente de surdité chez les plus de 50 ans, elle commence par une perte de distinction des sons aigus puis progressivement des sons graves.
La cochlée
La cochlée est un tube d’environ 32 mm de long et 2 mm de diamètre enroulée autour d’un axe creux qui contient le nerf auditif.
À la base de la cochlée se trouve deux trous recouvert d’une membrane : la fenêtre ovale, localisée sous la partie plate de l’étrier, et la fenêtre ronde.
Sur une coupe transversale de la cochlée, On peut voir que le tube est divisé en trois compartiments nommés « rampes », remplis d’un fluide ; il y a la rampe vestibulaire, la rampe tympanique, et le canal cochléaire.
Image 4
Les trois canaux parallèles de la cochlée

La périlymphe est le liquide présent dans la rampe vestibulaire et dans la rampe tympanique. La composition de la périlymphe est similaire à celle du liquide céphalorachidien : elle a une faible concentration en ions potassium K+ (7 mM) et une forte concentration en ions sodium Na+ (140 mM).
Le canal cochléaire est rempli d’endolymphe dont les concentrations ioniques sont semblables à celles du milieu intracellulaire, il y a une forte concentration en ions potassium K + (150 mM) et une faible concentration du sodium Na+ (1 mM), pourtant c’est un milieu extracellulaire.
On peut expliquer cette différence de concentration par l’activation d’un mécanisme de transport actif de la stria vascularis. La stria vascularis réabsorbe le sodium de l’endolymphe et sécrète du potassium dans l’endolymphe contre leurs gradients de concentration.
L’endolymphe présente ainsi un potentiel électrique d’environ 80 mV plus positif que celui de la périlymphe en raison de ces différences de concentration ionique et de la perméabilité de la membrane de Reissner. On appelle ceci le potentiel endocochléaire, qui représente un facteur important favorisant la transduction auditive.
Les trois rampes s’enroulent en spirale à l’intérieur de la cochlée. La rampe vestibulaire et le canal cochléaire sont séparés par la membrane de Reissner. La membrane basilaire qui supporte l’organe de Corti sépare la rampe tympanique et le canal cochléaire.
La rampe tympanique se termine par la fenêtre ronde et la rampe vestibulaire est adjacente à la fenêtre ovale.
Les deux rampes communiquent au sommet de la cochlée appelé apex dans une région appelée hélicotrème (= ouverture de la spirale). Le fluide de la rampe vestibulaire se trouve donc en continuité avec celui de la rampe tympanique.
Physiologie de la cochlée
Tout mouvement de la membrane de la fenêtre ovale doit être associé un mouvement complémentaire de la membrane de la fenêtre ronde. Ce mouvement est inévitable car la cochlée est remplie d’un milieu liquide incompressible se trouvant dans une cavité osseuse rigide.
Ainsi on peut comparer la pression s’exerçant sur la membrane de la fenêtre ovale à la pression que l’on exerce sur l’extrémité d’un ballon lorsqu’il est rempli d’eau. En effet, dans ce cas c’est l’autre extrémité du ballon qui se déforme. Cependant, il faut garder en tête que les structures à l’intérieur de la cochlée ne sont pas rigide, donc cet exemple ne décrit pas exactement la réalité.
En outre, la membrane basilaire est souple et en présence d’ondes sonores, elles s’infléchit. Elle monte et descend et fait à son tour osciller la partie basale du conduit cochléaire. Selon la fréquence des sons envoyés à l’oreille (aigus ou graves), ce n’est pas le même endroit sur la membrane basilaire qui va vibrer. Les sons aigus font vibrer la membrane basilaire proche de la fenêtre ovale, tandis que les sons graves la font vibrer vers l’apex.
La membrane basilaire est sensible aux sons. Cette sensibilité se détermine par deux propriétés structurales. Dans un premier temps, la membrane est plus large à l’apex qu’à la base, d’un facteur cinq environ. Dans un deuxième temps, la rigidité de la membrane diminue de la base à l’apex, la base étant un peu près 100 fois plus rigide. Cette membrane peut être comparé à la palme des nageurs dont la base est étroite et ferme, et l’extrémité large et souple.La chaîne des osselets transfère les pressions acoustiques du milieu aérien au milieu liquidien de la cochlée. L’étrier vibre d’avant en arrière ce qui entraîne des vibrations dans les liquides qui vont arriver à faire vibrer les différentes membranes. C’est en repoussant la partie plate de l’étrier contre la fenêtre ovale que le son entraîne un déplacement de la périlymphe dans la rampe vestibulaire mais aussi de l’endolymphe dans le canal cochléaire.
Ainsi, la membrane basilaire s’élargit en allant vers l’apex tandis que la cochlée se rétrécit progressivement de la base jusqu’à l’apex.
Image 5
Membrane basiliaire représentée dans une cochlée déroulée

Sur la membrane basilaire sont disposées les cellules nerveuses (les cellules ciliées) du récepteur auditif qui détectent les vibrations et vont envoyer les messages vers le cerveau. La paroi basilaire du canal cochléaire est la plus épaisse car elle renferme l’organe sensoriel auditif, ou organe de Corti.
LE SAVIEZ-VOUS ?
Un implant cochléaire est un dispositif médical électronique qui remplace les fonctions endommagées de l’oreille interne. Contrairement aux prothèses auditives qui amplifient les sons, les implants cochléaires remplissent les fonctions normalement assurées par les parties endommagées de l’oreille interne, et plus précisément de la cochlée afin de fournir au cerveau des signaux sonores.
L’organe de Corti et les cellules ciliées
L’organe de Corti se situe dans l’oreille interne, et plus précisément dans le canal de la cochlée. Il abrite les récepteurs auditifs, c’est l’organe de la perception auditive. Au dessus de l’organe de Corti se situe la membrane tectoriale. Cette membrane est l’élément sensible de l’ouïe. Elle comprend environ 14000 cellules ciliées au contact desquelles prennent naissance les fibres du nerf auditif.
L’organe de Corti présente deux types de mécanorécepteurs. D’une part il y a les cellules ciliées internes qui sont disposées sur une seule rangée et qui constituent le récepteur sensoriel. Elles sont chacune sensibles à une certaine fréquence et assurent un rôle de transduction faisant converger l’information vers le nerf auditif qui est une voie afférente en direction des centres nerveux. D’autre part, il y a les cellules ciliées externes disposées sur trois rangées et qui possèdent un système contractile qui permet d’amplifier une vibration locale : c’est l’électromotilité. Le nombre de cellules ciliées externes est largement supérieur à celui des cellules ciliées internes.
Image 6
Les cellules ciliées de l’organe de Corti

Arrangement des cellules ciliées à la base de la cochlée en vue de surface en microscopie électronique à balayage (MEB)
Image 7

On peut observer l’organisation très régulière des cellules ciliées et de leurs stéréocils très courts. Les stéréocils sont composés de filaments d’actine.
La disposition en W des cils des cellules ciliées externes, fermement ancrés dans la membrane tectoriale est à corréler avec les mécanismes actifs nécessaires à la physiologie des hautes fréquences.
Lorsque l’étrier fait vibrer la périlymphe, les vibrations font onduler la membrane basilaire. Les cils des cellules ciliées externes se déplacent horizontalement ce qui entraînent la dépolarisation de ces cellules et leur contraction. Les cellules ciliées externes vont ainsi tirer périodiquement sur la membrane tectoriale et donc la mettre en contact avec les cils des cellules ciliées internes. Ces derniers se couchent et provoquent la dépolarisation des cellules ciliées internes. Celles-ci libèrent alors du glutamate dans l’espace intersynaptique avec les fibres du nerf auditif. L’organe de Corti est aussi capable de moduler le signal auditif. Les cellules ciliées externes peuvent amplifier le signal par un processus appelé électromotilité. Il augmente le mouvement de la membrane basilaire et donc augmente la déviation des stéréocils dans les cellules ciliées internes.
La protéine Prestin est un élément crucial de cette amplification cochléaire qui change de forme en fonction du niveau de tension à l’intérieur de la cellule ciliée externe. Lorsque la cellule ciliée externe est dépolarisée la protéine Prestin se raccourcit, et parce qu’elle se trouve sur la membrane des cellules ciliées externes, elle tire sur la membrane basilaire. Ainsi, cela crée un effet plus intense sur les cellules ciliées internes. A l’inverse, lorsque la cellule ciliée externe est hyperpolarisée, la protéine Prestin allonge et soulage la tension sur les cellules ciliées internes, ce qui diminue les impulsions nerveuses au cerveau. Ainsi, les cellules ciliées sont capables de modifier le signal sonore avant d’atteindre le cerveau.
LE SAVIEZ-VOUS ?
Lors d’un traumatisme sonore important, de la prise de médicaments toxiques ou d’excitotoxicité, c’est-à-dire qu’il y a de l’acide glutamique en excès, les terminaisons post-synaptiques des cellules ciliées internes peuvent gonfler et exploser. Cela peut entraîner ainsi une surdité totale. Si le corps cellulaire n’est pas atteint, le processus est réversible. Dans l’autre cas, l’individu devient sourd.
Image 8
L’organe de Corti

Ce schéma nous permet d’observer que les cellules ciliées internes, les cellules ciliées externes et les piliers de Corti sont insérés entre la membrane basilaire est une mince couche de tissu, appelée lame réticulaire. De même, nous pouvons voir la membrane tectoriale qui recouvre les stéréocils des cellules ciliées. Par ailleurs, les piliers de Corti se dressent d’une membrane à l’autre, et forment un support.
Réception du signal sonore par les cellules ciliées
Ces cellules ciliées se déploient sur quatre rangées. Les cellules ciliées située entre l’axe de la cochlée et les piliers de Corti, sont des cellules ciliées internes. Elles sont au nombre de 4500 environ, disposer sur un seul rang. Les cellules situées au-delà des piliers sont dénommées cellules ciliées externes. Elles sont environ 12 000 à 20 000 cellules chez l’homme, disposées en trois rangées.
Ces rangées s’étendent sur toute la longueur de la membrane basilaire de la base au sommet, c’est-à-dire, l’apex de la cochlée.
On qualifie ces cellules de “ciliées” car leur pôle apical en contact avec l’endolymphe porte une centaine de stéréocils.
Sous l’effet du son, la fenêtre ovale bouge, faisant se déplacer la membrane basilaire: les cellules ciliées internes, solidaires de la membrane basilaire s’inclinent en rentrant en contact avec la membrane tectoriale au travers de laquelle transitent les informations destinées au cerveau. Cependant, ce mécanisme ne fonctionne que pour un niveau sonore supérieur à 50 dB.
Pour des sons inférieurs à 50 dB, la membrane basilaire se déforme mais pas suffisamment pour incliner les cellules ciliées internes. Ce sont alors les cellules ciliées externes qui se contractent. En effet, elles sont munies de mécanismes contractiles actifs, contrairement aux cellules ciliées internes. Cela a pour effet de déplacer la membrane tectoriale, elle entre alors en contact avec les cellules ciliées internes, qui permettent ainsi la transmission des informations au cerveau pour des sons inférieurs à 50 dB.
L’étirement des cils des cellules ciliées internes provoque un mouvement ionique donc une modification du potentiel transmembranaire et la libération d’un neurotransmetteur : le message est envoyé au système nerveux. Ce mécanisme fait parti de la transduction, il est détaillé dans l’onglet “transmission”.
Lorsqu’un signal sonore pénètre dans l’oreille, cela engendre la déviation des stéréocils de la cellule ciliée interne ce qui ouvre mécaniquement des canaux ioniques, qui permettent l’entrée d’ions K+ dans la cellule ciliée interne. Cet afflux d’ions positifs, venant de l’endolymphe, dépolarise la cellule et entraîne un potentiel de récepteur. Ce potentiel de récepteur ouvre des canaux calciques voltage-dépendants, et des ions calcium Ca2+ pénètrent dans la cellule. Cette entrée d’ions-calcium déclenche la libération d’un neurotransmetteur à la base de la cellule : le glutamate. La libération du glutamate est réalisée grâce à des structures particulières fixées à la base de la cellule ciliée interne, ce sont : les rubans synaptiques.
À chaque cellule ciliée interne est connectée au moins une dizaine de fibres afférentes. Ces fibres sont de trois catégories : les fibres à bas seuil, qui sont concernent les sons faibles, les fibres à seuil modéré qui concernent les sons modérés et les fibres à haut seuil qui concernent les sons forts. Chacune des fibres nerveuses afférentes est reliée à un ruban synaptique d’une cellule ciliée interne.
Le glutamate se fixe sur des récepteurs spécifiques de fibres nerveuses afférentes.
Ainsi, les cellules ciliées internes sont des cellules vibratiles, dont la mise en mouvement par une onde sonore déclenche la libération de neurotransmetteurs. Ces cellules sont donc des « capteurs de vibrations », et sont connectées à la voie afférente.
D’autre part, les cellules ciliées externes sont des cellules contractiles activées par le système efférent. C’est-à-dire que la « contraction » est commandée par le système nerveux. Elles jouent un rôle essentiel dans la sélectivité fréquentielle. Autrement dit, dans la capacité d’un individu à distinguer les unes des autres des fréquences sensiblement identiques jouées en même temps.
LE SAVIEZ-VOUS ?
Pratiquement inexistante chez le poisson, l’audition est remplacée par une sensibilité aux vibrations. C’est la ligne latérale qui permet de percevoir ces oscillations, c’est l’appareil sensoriel du poisson. La ligne latérale est une série de petits points régulièrement espacés placés sur les flancs du poisson qui sont reliés au système nerveux. Elle permet au poisson de se renseigner sur l’eau ambiante, c’est-à-dire sur le courant, la profondeur, la température et de percevoir les vibrations comme les chocs, les battements et mouvements de nage.
Synthèse du fonctionnement cochléaire
Pour entendre un son, il faut que l’oreille reçoive un signal sonore, ce qui engendrera une stimulation sonore dans la cochlée. Voici, de manière synthétique comment l’oreille réceptionne le signal sonore.
La vibration liquidienne se transmet à la membrane basilaire, ainsi qu’à toute la cloison cochléaire qui se déplace vers le haut, c’est-à-dire, vers la rampe vestibulaire et vers le bas, c’est-à-dire, vers la rampe tympanique. Cette vibration respecte une tonotopie grossière liée aux propriétés élastiques de la cloison cochléaire.
Ensuite, lorsque la membrane basilaire s’incurve vers la rampe vestibulaire, la membrane tectoriale effectue un mouvement de glissement relatif vers l’extérieur entraînant le cisaillement des cils les plus longs des cellules ciliées externes et leur déplacement dans la direction excitatrice. Les cellules ciliées externes sont bio-électriquement activées. Cela signifie qu’il y a une variation de leur potentiel membranaire.
Les cellules ciliées externes excitées se contractent en phase avec la fréquence stimulante. Le couplage qu’elles réalisent entre la membrane tectoriale entraîne une amplification de la vibration dans la région très restreinte de la cloison cochléaire.
Dans la zone étroite où s’est focalisée l’énergie libérée par le mécanisme actif, un petit nombre de cellules ciliées internes, voire une seule, est excité avec un maximum de sensibilité.
Les cellules ciliées internes dépolarisées libèrent leur neurotransmetteur et le message est envoyé au système nerveux central par les fibres afférentes avec qui elles font synapse.
En résumé, les cellules ciliées internes sont des récepteurs sensoriels passifs qui transforment l’énergie vibratoire qui leur est transmise par la vibration de la membrane basilaire, amplifiée par la contraction active des cellules ciliées externes. Les mécanismes actifs sous-tendus par les cellules ciliées externes sont donc responsables de la grande sensibilité et sélectivité en fréquence retrouvées au niveau de la membrane basilaire, des cellules ciliées internes et des fibres du nerf auditif de la cochlée.
Image 8
Coupe transversale détaillée d’un tour du canal cochléaire.
Les ondes sonores engendrent le déplacement dans des directions opposées des fenêtres ovales et rondes situées à la base de la cochlée. Cela fait que la membrane basilaire se déplace et déclenche une onde progressive qui balaye de la base vers le sommet de la cochlée. L’amplitude de l’onde progressive augmente à mesure qu’elle se déplace et atteint un pic à un endroit directement lié à la fréquence du son.

La figure 8 illustre un grossissement de l’organe de Corti. L’onde progressive fait monter et descendre la membrane basilaire et donc l’organe de Corti. L’organe de Corti possède un renforcement central formé de piliers. Les cellules ciliées sont au sommet de l’organe de Corti. La membrane tectoriale est maintenue en place par un mécanisme semblable à une charnière du côté de l’organe de Corti et flotte au-dessus des cellules ciliées. Lorsque les membranes basilaires et tectoriale se déplacent avec l’onde, le mécanisme de charnière provoque le déplacement latéral de la membrane tectoriale au-dessus des cellules ciliées. Ce mouvement de cisaillement latéral plie les cils au sommet des cellules ciliées, cela tire sur les maillons fins et ouvre les canaux de la trappe. Il y a un afflux de potassium, puis de calcium, ce qui provoque la libération de neurotransmetteurs. Ce mécanisme est étudié plus en détail dans la partie transmission.
LE SAVIEZ-VOUS ?
Les chiens ont une oreille sensible qui leur permet de percevoir des sons inaudibles par l’homme. Les ultrasons sont des sons très aigus qui provoquent une gêne, mais pas de douleur. Ils sont utilisés pour dresser un animal, maîtriser les aboiements ou encore éloigner un chien ou un chat intrus.
